Kira-kira 40 persen dari seluruh tubuh terdiri dari otot rangka dan kira- kira 10 persen lainnya terdiri dari otot jantung dan otot polos. Otot rangka dibentuk oleh berbagai jenis jaringan, jaringan-jaringan ini terdiri atas jaringan saraf, pembuluh darah, jaringan ikat, dan sejumlah serat otot sendiri yang diameternya berkisar dari 10 sampai 80 mikrometer, masing-masing serat ini terbuat dari rangkaian subunit yang lebih kecil.
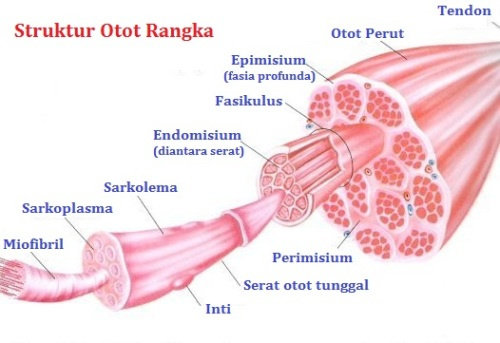
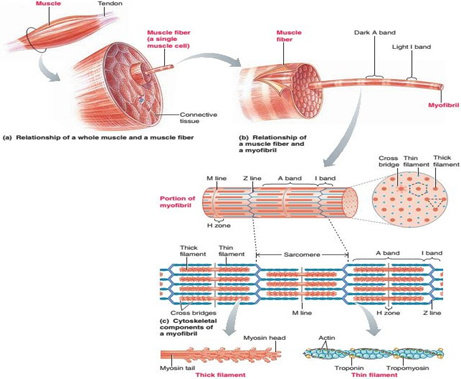
Terdapat tiga lapisan jaringan ikat dalam serabut otot rangka, lapisan terluar yang melapisi seluruh otot disebut epimisium, di dalam lapisan ini terdapat lapisan perimisium yaitu lapisan jaringan ikat yang membungkus satu kelompok serabut otot tersendiri yang disebut fasikuli. Masing-masing serabut otot di dalam fasikuli dibungkus oleh jaringan ikat yang disebut endomisium.
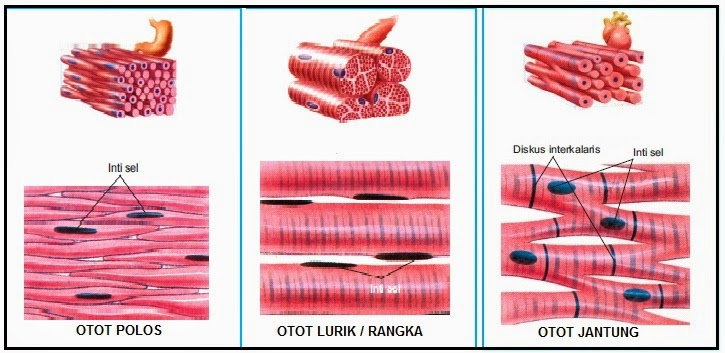
Sel-sel otot memiliki bentuk yang unik, walau demikian sel-sel ini memiliki organela-organela yang hampir sama dengan yang dimiliki sel lain pada umumnya, seperti mitokondria, lisosom, dan lainnya. Namun, tidak seperti kebanyakan sel dalam tubuh, sel-sel otot memiliki inti yang multinuclear atau lebih dari satu. Salah satu ciri khas lain dari sel ini ialah penamakan garis garis striae, garis ini dihasilkan dari pergantian bagian gelap dan terang di sepanjang serabut otot.
Gambar Struktur Otot Rangka
Sarkolema adalah membran serabut otot yang terdiri dari membran sel yang sebenarnya, yang disebut membran plasma dan sebuah lapisan luar yang terdiri dari satu lapisan tipis bahan polisakarida yang mengandung sejumlah serat kolagen tipis. Pada ujung serabut otot, lapisan permukaan sarkolema ini bersatu dengan serat tendon dan kemudian serat-serat tendon berkumpul menjadi berkas untuk membentuk tendon otot dan kemudian menyisip ke dalam tulang. Di dalam sarkolema terdapatlah sarkoplasma.
Sarkoplasma adalah matriks yang terdiri dari unsur-unsur intraselular yang di dalamnya mengandung protein-protein selular, organela, dan miofibril. Miofibril merupakan struktur threadlike berjumlah banyak yang mengandung protein kontraktil. Miofibril ini disusun oleh dua tipe filamen protein yaitu, filamen tebal yang disusun oleh protein miosin dan filamen tipis yang disusun oleh protein aktin. Di dalam aktin terdapat tambahan protein yaitu troponin dan tropomiosin, molekul ini berukuran kecil di dalam otot, namun memegang peranan penting dalam regulasi proses kontraksi otot. Letak dari filamen filamen inilah yang mengakibatkan terbentuknya penampakan striae pada serabut-serabut otot.
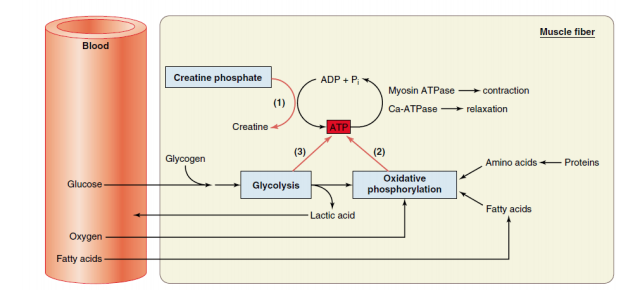
Miofibril kemudian dapat dibagi lagi ke dalam segmen-segmen tersendiri yang disebut sebagai sarkomer. Sarkomer ini kemudian dipisahkan satu dengan lainnya oleh selaput tipis protein struktural yang disebut garis Z. Filamen miosin terletak terutama di daerah gelap dari sarkomer, bagian ini dinamakan sebagai pita A, sedangkan filamen aktin terletak terutama di daerah terang dari sarkomer, bagian ini dinamakan sebagai pita I. Pada pertengahan sarkomer, terdapat bagian dari filamen miosin yang tidak saling bertumpang tindih dengan aktin, bagian ini disebut sebaga zona H. Cairan sarkoplasma mengandung kalium, magnesium, fosfat, enzim protein dalam jumlah besar, dan mitokondria dalam jumlah yang banyak yang terletak di antara dan sejajar dengan miofibril. Terdapatnya mitokondria dalam jumlah yang banyak serta terletak di antara dan sejajar dengan miofibril menunjukkan bahwa miofibril-miofibril yang berkontraksi membutuhkan sejumlah besar adenosin trifosfat (ATP) yang dibentuk oleh mitokondria.
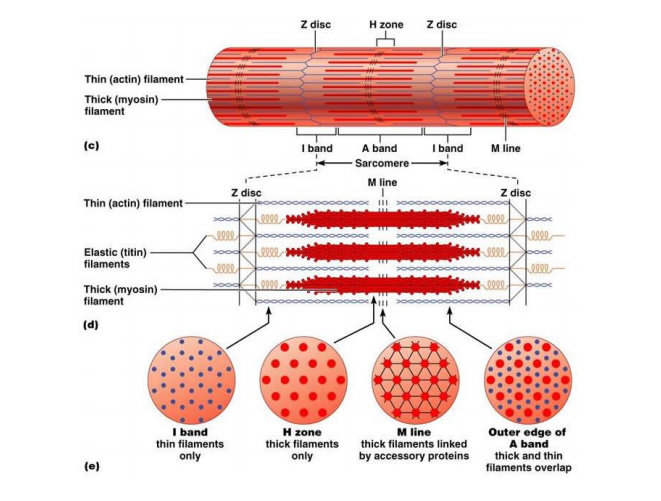
Gambar Struktur Sarkomer
Retikulum sarkoplasmik merupakan nama lain dari retikulum endoplasma yang terdapat di dalam sarkoplasma serat otot. Struktur ini penting untuk menimbulkan kontraksi otot yang cepat, semakin cepat kontraksi suatu otot, maka ia mempunyai banyak sekali retikulum sarkoplasmik.5
Kontraksi Otot
Potensial aksi tunggal menyebabkan kontraksi singkat yang kemudian diikuti dengan relaksasi. Respon ini disebut sebagai kontraksi kedutan otot (muscle twitch). Kedutan timbul kira-kira 2 mdet setelah dimulainya depolarisasi membrane, sebelum repolarisasi selesai. Lamanya kontraksi kedutan beragam, sesuai dengan jenis serabut otot yang dirangsang. Serabut otot cepat, yang terutama berperan dalam gerakan halus, cepat dan tepat, mempunyai lama kedutan 7,5 mdet. Serabut otot lambat, yang terutama berperan dalam gerakan kuat, menyeluruh, dan dipertahankan, memiliki lama kedutan sampai 100 mdet.
Dasar Molekular Kontraksi
Proses yang mendasari pemendekan elemen kontraktil di otot adalah pergeseran filamen tipis pada filamen tebal. Lebar pita A tetap, sedangkan garis Z bergerak saling mendekat ketika otot berkontraksi dan saling menjauh bila otot diregang.
Pergeseran selama kontraksi otot terjadi bila kepala miosin berikatan erat dengan aktin, menekuk di taut kepala dengan leher, dan kemudian terlepas. Lonjakan tenaga (power stroke) ini bergantung pada hidrolisis ATP yang serentak. Molekul miosin-II adalah dimer yang memiliki dua kepala, tetapi setiap saat hanya satu yang melekat ke aktin.
Banyak kepala miosin mengalami siklus pada saat yang sama atau hampir bersamaan, dan kepala- kepala tersebut bersiklus berulang-ulang untuk menghasilkan kontraksi otot keseluruhan. Setiap power stroke akan memendekkan sarkomer sekitar 10 nm. Setiap filamen tebal mengandung 500 kepala miosin, dan setiap kepala bersiklus sekitar lima kali per detik selama berlangsungnya kontraksi cepat.
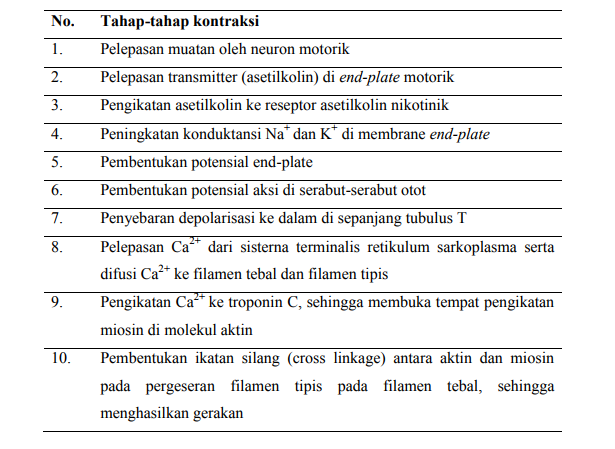
Proses ketika depolarisasi serabut otot memicu kontraksi disebut dengan penggabungan eksitasi-kontraksi (excitation-contraction coupling). Ururtan peristiwa yang berperan dalam kontraksi dan relaksasi otot rangka dirangkum pada tabel di bawah:
Tabel Tahap Kontraksi Otot
Tabel Tahap Relaksasi Otot

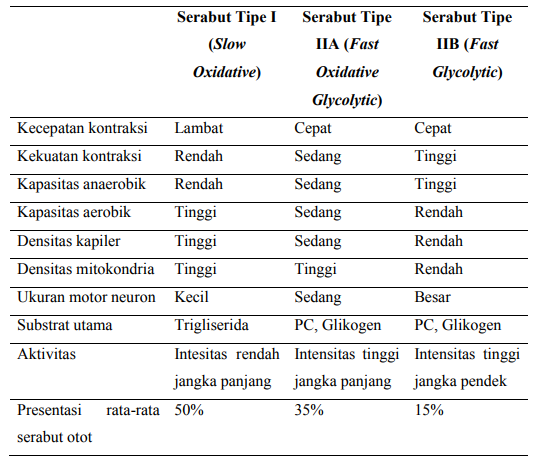
Tipe Serabut Otot
Serat-serat otot terdiri atas dua jenis serabut, yaitu serabut otot tipe I, serabut lambat, serabut merah, atau serabut oksidatif lambat (slow-twitch muscle fiber) dan serabut otot tipe II, serabut cepat, serabut putih, atau serabut otot anaerobik (fast-twitch muscle fiber).
Pada serabut tipe II masih dibagi menjadi dua macam, yaitu tipe IIa dan tipe IIb. Sehingga dapat diklasifikasikan menjadi 3 jenis serabut otot, yaitu tipe I (slow twitch oxidative), tipe IIa (fast twitch oxidative), dan tipe IIb (fast twitch glycolytic).
Serabut otot tipe lambat
Serabut otot tipe lambat mengandung enzim oksidatif dalam jumlah yang besar, berkontraksi secara lambat dan melepaskan energi secara bertahap sesuai dengan kebutuhan yang diperlukan oleh tubuh pada keadaan aktivitas steady-state misalnya joging, bersepeda, dan endurance swimming.
Serabut- serabut otot ini mengandung lebih banyak mitokondria, suplai pembuluh darah, dan mioglobin sehingga dapat secara efisien dalam menggunakan oksigen untuk menghasilkan energi, membuatnya resisten terhadap kelelahan namun tidak dapat menghasilkan energi atau daya sebagaimana serabut otot tipe cepat.
Mioglobin yang lebih banyak terkumpul di dalam serabut tipe ini menyebabkan warna serabut ini menjadi lebih merah, karena mengandung pigmen mioglobin (seperti hemoglobin) di dalamnya. Ketika tubuh melakukan aktivitas tipe ketahanan, maka serabut otot tipe lambat ini akan lebih banyak digunakan untuk pergerakan sebab serabut otot jenis ini akan memenuhi kebutuhan energi dari otot yang bekerja.
Serabut otot tipe cepat
Sedangkan serabut otot tipe cepat berkontraksi secara cepat dan melepaskan energi secara cepat, hal ini disebabkan serabut otot tipe ini mengandung lebih banyak retikulum sarkoplasma sehingga lebih cepat dalam melepaskan dan mengambil kembalik ion kalsium, struktur kepala myosin juga sedikit berbeda dibanding serabut otot tipe lambat menyebabkan serabut otot ini lebih efisien dalam menghidrolisa ATP, namun serabut otot ini rentan terhadap kelelahan yang disebabkan jalur penghasil energi yang digunakan yaitu sistem metabolisme anaerobik.
Tubuh banyak menggunakan serabut otot jenis ini untuk melakukan tipe aktivitas daya ledak, seperti angkat beban, senam atletik, dan lari sprint. Pada setiap individu rasio antara serabut otot tipe lambat dan cepat berbeda dan telah dideterminasi secara genetik, sehingga dapat menjadikan mereka lebih cocok pada suatu cabang olahraga atau aktivitas tertentu.
Latihan fisik yang tepat akan turut mengembangkan dan menimbulkan adaptasi yang tepat bagi tiap tipe serabut otot.
Serabut otot tipe cepat akan menunjukkan perkembangan dan manfaat dengan latihan anaerobik, seperti misalnya lari sprint atau latihan dengan interval dan latihan beban.
Serabut otot tipe lambat akan menunjukkan perkembangan dan manfaat terutama dari aktivitas ketahanan yang menggunakan jalur sistem aerobik, seperti berlari, bersepeda, dan berenang.
Tabel Tipe Serabut Otot