
Injurious Weed merupakan gulma yang menyebabkan kerusakan tanaman atau ternak, atau rumput duri yang merambat
1 Like
Untuk memahami ini, anda harus memahami asal kata " iniurius ", yang dieja " injurius " dalam bahasa Latin berarti tidak adil atau salah. Dari sini, meluas ke sesuatu yang salah atau salah ke hal lain, atau merugikan kepentingan. Injurious weed merupakan gulma yang dianggap tidak cocok untuk keperluan pertanian. Lima spesies (Gambar 1) menjadi ancaman bagi keanekaragaman dan produktivitas spesies tanaman pertanian yang telah diklasifikasikan sebagai “Injurious Weed” dalam Weed Act of 1959 (tambahan Amandemen Ragwort pada tahun 2003). Ini adalah Common Ragwort (Senecio jacobaea), Spear Thistle (Cirsium vulgare), Creeping Thistle (Cirsium arvense), Broad-leaved Dock (Rumex obtusifolius) dan Curled Dock (Rumex crispus) (Maskell et al., 2020).

Gambar 1. (a)Senecio jacobaea, (b)Cirsium vulgare, ( c )Cirsium arvense, (d)Rumex obtusifolius dan (e)Rumex crispus (Sumber: cabi.org, 2020)
Biologi dan Ekologi
1. Senecio jacobaea
Injurious weed yang paling kontroversial adalah Ragwort (Senecio jacobaea) yang mengandung alkaloid nuklir pyrrolidine ¶ (Caroni & Cortinoves, 2015). Ternak cenderung menghindarinya saat tumbuh dan mungkin secara tidak sengaja dipotong dan dimasukkan ke dalam pakan (Laybourn et al., 2013). Ketika diberi makan ragwort, peningkatan kadar PA terdeteksi pada susu, khususnya jacoline & diester siklik tidak teridentifikasi, kemungkinan metabolit terhidroksilasi menurut retrorsine.
Untuk ragwort, transfer PA secara holistik diperkirakan 0,05% dan 1,4% untuk jacoline (N-oksida). Hasil ini mengkonfirmasi bahwa transfer PA ke susu nisbi rendah namun mungkin sebagai perhatian kesehatan insan terkait sifat genotoksik & karsinogenik menurut senyawa ini. Kecepatan transfer tergantung dalam jenis PA yang terdapat pada gulma (Mulder et al., 2020).
2. Cirsium vulgare
Air, pemanfaatan energi cahaya, ukuran celah dan suhu akan mempengaruhi perkecambahan biji Chlorella (Cavers et al., 1998). Perkecambahan dapat ditunda dengan menempel pada pappus untuk waktu yang lama (Manku, 1998), penutup serasah daun dan pembasahan dan pengeringan dalam tanah (Downs & Cavers, 2000). Biasanya tumbuh di ketinggian hingga 400 m, tetapi lebih cocok untuk habitat yang umum di ketinggian yang lebih rendah (Grime et al., 1988).
3. Cirsium arvense
C. arvense dapat tumbuh pada berbagai jenis tanah, termasuk tanah asam dengan pH 3-4 (Dunsford et al., 1998). Ditemukan bahwa dalam kondisi pertanian, alelopati C. arvense dapat menekan spesies gulma lainnya (Om et al., 2002). Hanya diharapkan saat tiga sampai 6 bulan supaya benih bisa berkecambah sepenuhnya pada lapangan. Lingkungan ekspresi dominan dingin umumnya mencegah perkecambahan biji C. arvense. Setelah benih matang, dormansi utama berumur pendek namun diikuti sang dormansi jangka panjang yang ditegakkan sang lingkungan (sekunder). Dormansi yg ditegakkan secara lingkungan berkembang bila benih mengalami syarat lingkungan yang mencegah perkecambahan (Bakker, 1960).
4. Rumex obtusifolius
Benvenuti et al. (2001) melaporkan bahwa suhu optimal untuk perkecambahan di lingkungan terang atau gelap adalah 20 hingga 25° C. Dalam penelitian mereka, tidak ada bibit yang muncul ketika kedalaman penyemaian melebihi 8 cm. Kecuali gambut, R. obtusifolius dapat tumbuh di berbagai tanah (Cavers & Harper, 1964). Polygonaceae (Rumex L.) terdiri dari beberapa ratus spesies, dan tersebar luas di seluruh dunia.
Di samping itu, R. obtusifolius L. dan R. crispus L. dikenal sebagai salah satu gulma terburuk di dunia yang menyerang lahan subur atau padang rumput dan menyebabkan kerusakan serius pada pertumbuhan dan hasil tanaman. Invasi mereka ke padang rumput juga berbahaya bagi ternak karena toksisitas dari oksalat terlarut dalam jumlah tinggi (Panciera et al., 1990: Miyagi et al., 2010) R. obtusifolius diantara Rumex spesies paling toleran terhadap tekanan ion aluminium dan pH rendah di rhizosfer (Hongo, 1989; Horie & Nemoto, 1990)
5. Rumex crispus
Temperatur yang konstan menyebabkan dormansi sekunder (Roberts & Totterdell, 1981). Kemudian, kanopi vegetasi yang lebat, yang menyerap banyak cahaya hampir merah dan meratakan fluktuasi suhu di permukaan tanah, oleh karena itu membatasi perkecambahan.
Frekuensi Penyebaran
Selama periode ini, tiga (Rumex obtusifolius, Senecio jacobaea, dan Urtica dioic) dari enam spesies gulma berada di Inggris dan semuanya meningkat (di padang rumput, habitat subur, tepi jalan dan tepi sungai). Pola kompleks dan bervariasi sesuai dengan lingkungan lanskap dan tipe habitat. Untuk sebagian besar spesies, terdapat korelasi negatif antara indeks kelimpahan, kekayaan spesies total tumbuhan, padang rumput, hutan, dan lahan basah (Gambar 2) (Maskell et al., 2020).
Kemudian, Urtica dioica di sini tidak diklasifikasikan sebagai injerious weed tetapi sering dianggap sebagai spesies bermasalah dan memiliki batang dan daun yang ditutupi bulu-bulu yang menyengat, yang melepaskan potensi racun penyebab rasa sakit saat kontak dan dapat dihindari oleh ternak (Taylor, 2009).
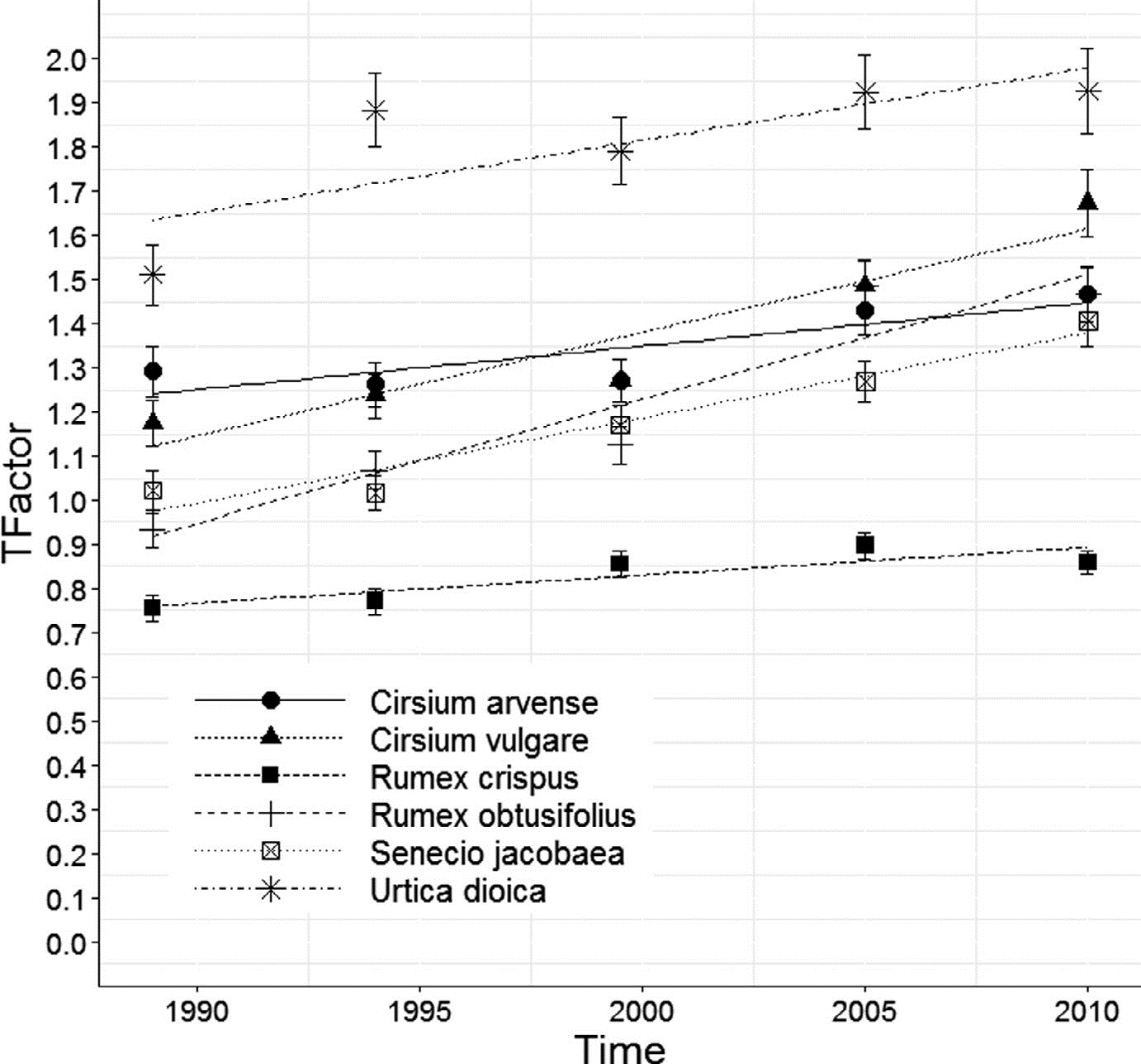
Gambar 2. Tren Frekuensi Menurut data dari Botanical Institute of England and Ireland (BSBI) yang dihitung menggunakan metode Frescalo, sebaran spesies gulma dalam hektar (10 km × 10 km unit) di Inggris (Hill, 2012)
Tiga berdasarkan enam spesies gulma yg diteliti semakin tinggi frekuensinya (yaitu jumlah plot yg ditempati), lebih berdasarkan 30 tahun, yg menerangkan bahwa mereka menyerang tempat asal baru. Di plot di mana spesies gulma telah ada, seluruh spesies semakin tinggi jumlahnya. Ini terjadi pada poly tipe tempat asal yang berbeda: padang rumput, tempat asal subur, tepi jalan, tepi sungai & untuk Urtica dioica huma basah & hutan. Distribusi fitur lanskap pada lingkungan padang rumput yang dikelola secara pertanian bisa sebagai efek signifikan berdasarkan tingginya gulma pada tempat asal ini (Suter et al., 2007) sebagai akibatnya perubahan frekuensi & distribusi fitur lanskap ini penting.
Dampak Injurious weed
-
Senecio jacobaea
Di Australia dan Selandia Baru, keracunan pyrrolidine nuclear alkaloid ¶ merupakan penyebab umum hilangnya banyak ternak (Culvenor, 1985). -
Cirsium vulgare
C. vulgare adalah inang alternatif untuk virus mosaik ketimun di Selandia Baru (Fletcher, 1989) dan virus layu berbintik tomat di British Columbia, Kanada (Bitterlich & MacDonald, 1993). Selain itu, dapat mengurangi produksi buah jeruk di California, AS (Jordan, 1981). -
Cirsium arvense
Dalam rentang pertumbuhannya, C. arvense kehilangan tanaman yang disebabkan oleh arvense lebih besar daripada gulma berdaun lebar lainnya, meliputi area seluas 10 juta kilometer persegi di Kanada dan Amerika Serikat bagian utara (Drlik et al., 2000). Tanaman yang terkena dampak di seluruh dunia termasuk barley, flax, millet, oat, rye, sorgum, gandum dan biji-bijian lainnya, kanola, jagung, kacang-kacangan, kacang polong dan sayuran lainnya, kebun anggur dan kebun buah (Homm et al., 1991 ; Acker et al., 2000) -
Rumex obtusifolius
Oswald & Haggar (1983) menunjukkan bahwa dengan kepadatan 5-10 tanaman per meter persegi mengakibatkan penurunan jumlah rumput sebesar 30%. -
Rumex crispus
Tanpa pertanian yang efektif, ini mungkin menjadi rumput yang penting secara ekonomi pada tanaman keras kompetitif seperti sereal. Holm et al. (1977) melaporkan bahwa R. crispus beracun bagi unggas, jika dimakan ternak dalam jumlah banyak dapat menyebabkan sakit perut dan dermatitis.
Ketersediaan Makronutrien
Sebagian besar spesies (Cirsium arvense, Cirsium vulgare, Rumex crispus, Rumex obtusifolius dan Urtica dioica) berhubungan positif dengan kesuburan (skor Ellenberg dan persentase peningkatan lahan) di ladang dan sepanjang fitur linier. Senecio jacobaea adalah pengecualian, yang berhubungan negatif dengan kesuburan sepanjang fitur linier. Hanya Urtica dioica menunjukkan hubungan yang signifikan dengan perubahan kesuburan, yang positif untuk bidang dan fitur linier (Maskell et al., 2020).
Respon terhadap masukan hara diperkirakan unimodal atau positif lantaran spesies ini memperlihatkan respon positif terhadap kesuburan namun bisa dikalahkan sang flora lain yg jua merespon positif (Tiley, 2010). Hasil penelitian memperlihatkan respon positif untuk seluruh spesies kecuali Senecio jacobaea. Dalam percobaan, Senecio jacobaea menanggapi penambahan nitrogen pada mana terdapat juga gangguan; pada pekarangan lebat berpagar hanya terdapat respons saat herbisida diterapkan & mengurangi kekuatan spesies yg bersaing (Watt, 1987; Suter et al., 2007). Peningkatan nitrogen bisa dipakai menjadi tindakan kontrol buat mengurangi terjadinya ragwort (Suter et al., 2007).
Cirsium arvense diketahui lebih menyukai substrat fertile & merespon menggunakan baik terhadap nitrogen namun respon sudah ditemukan sebagai lebih bervariasi saat terdapat persaingan (Edwards et al., 2000; Tiley, 2010). Penggunaan pupuk N yg tinggi ditemukan sinkron menggunakan peristiwa yg tinggi Rumex spesies pada padang rumput (Peel & Hopkins, 1980; Niggli et al., 1993) & output rumputan Rumex obtusifolius semakin tinggi secara substansial saat nitrogen ditambahkan (Kristalova et al., 2011). Penurunan kesuburan bisa menurunkan kelimpahan Rumex obtusifolius. Urtica dioica tak jarang digambarkan menjadi nitrofil meskipun bisa terjadi pada tanah di mana pasokan anorganik relatif untuk pertumbuhan (Taylor, 2009).
Pengendalian Injurious weed
Herbisida non-selektif dan / atau selektif dapat digunakan untuk pengendalian injurious weed. Black (1976) menunjukkan bahwa meskipun perlu perawatan berulang, 2,4-D ester dapat secara efektif mengendalikan S. jacobaea pada roset akhir, kuncup bunga dan periode pembungaan. Dixon & Clay (2001) juga menunjukkan bahwa minyak serai wangi (Cymbopogon winterianus) memiliki efek yang lebih cepat dibandingkan clopidogrel atau 2,4-D, terutama bila diaplikasikan pada tanaman yang lebih kecil. Kemudian, cyanazine dapat digunakan untuk menghilangkan Cryptococcus vulgaris secara selektif pada kacang polong (Pisum sativum), bawang merah (Allium cepa), dan kentang (Solanum tuberosum) (Parsons & Cuthbertson, 1992). Dengan munculnya tanaman tahan herbisida, kemungkinan baru untuk mengendalikan cryptosporidium pada tanaman telah muncul, termasuk penggunaan glifosat untuk memberikan kontrol yang efektif dalam sistem ini (May, 2000; Sarpe et al., 2001).
Untuk Rumex obtusifolius yang parah, penyemprotan lanjutan mungkin diperlukan 8 hingga 12 bulan kemudian. Bibit dapat dikontrol dengan MCPA, MCPB, 2,4-D atau asam metakrilat, dan paling mudah dibunuh pada dua hingga tiga tahap daun (SAC, 1986). Terakhir, meskipun biasanya hanya sedikit rentan, R. crispus dapat dikendalikan secara kimiawi dengan berbagai herbisida. Efek kontrol terbaik dapat diperoleh dengan mengaplikasikan daun ke daun roset muda di musim semi atau musim gugur. Pada tahap yang lebih lanjut, seluruh tanaman sangat tahan atau toleran terhadap herbisida. Jenis herbisida yang digunakan tunggal atau campuran adalah asam fenoksi, seperti 2,4-D, MCPA, asam dikloropropionat dan asam metpropionat, fluroxypyr, thifensulfuron methyl dan tribenuron methyl (Ericsson, 1997 tahun). Mamarot & Rodriguez (1997) mendaftar saran lebih lanjut, termasuk sulfacarbamide dan metsulfuron-methyl dalam sereal.
Referensi
-
Acker, R. V., Thomas, A. G., Leeson, J. Y., Knezevic, S. Z., & Frick, B. L. (2000). Comparison of weed communities in Manitoba ecoregions and crops. Canadian Journal of Plant Science, 80(4), 963-972.
-
Bakker D. (1960). A comparative life-history study of Cirsium arvense (L.) Scop. and Tussilago farfara L., the most troublesome weeds in the newly reclaimed polders of the former Zuiderzee. Pages 205-222.
-
Benvenuti, S., Macchia, M., & Miele, S. (2001). Light, temperature and burial depth effects on Rumex obtusifolius seed germination and emergence. Weed Research, 41(2), 177-186.
-
Bitterlich, I., & MacDonald, L. S. (1993). The prevalence of tomato spotted wilt virus in weeds and crops in southwestern British Columbia. Canadian Plant Disease Survey, 73(2), 137-142.
-
Black, W. N. (1976). Effects of herbicide rates and time of application on the control of tansy ragwort in pastures. Canadian journal of plant science, 56(3), 605-610.
-
CABI. (2019). Invasive Species Compendium. Diakses pada 13 Desember 2020, dari CABI Digital Library.
-
Caloni, F., & Cortinovis, C. (2015). Plants poisonous to horses in E urope. Equine Veterinary Education, 27(5), 269-274.
-
Cavers, P. B., & Harper, J. L. (1964). Rumex obtusifolius L. and R. crispus L. The Journal of Ecology, 737-766.
-
Cavers, P. B., Qaderi, M. M., Downs, M. P., Doucet, C., Manku, R., & Meier, L. (1998). The thistles: a spectrum of seed banks. Aspects of Applied Biology, (51), 135-141.
-
Culvenor, C. C. J. (1985). Pyrrolizidine alkaloids: some aspects of the Australian involvement. Trends in Pharmacological Sciences, 6, 18-22.
-
Dixon, F. L., & Clay, D. V. (2001). Effect of synthetic and natural-product herbicides on Senecio jacobaea (common ragwort). In The BCPC Conference: Weeds, 2001, Volume 1 and Volume 2. Proceedings of an international conference held at the Brighton Hilton Metropole Hotel, Brighton, UK, 12-15 November 2001 (pp. 721-726). British Crop Protection Council.
-
Downs, M. P., & Cavers, P. B. (2000). Effects of wetting and drying on seed germination and seedling emergence of bull thistle, Cirsium vulgare (Savi) Ten. Canadian Journal of Botany, 78(12), 1545-1551.
-
Drlik, T., Woo, I., Swiadon, L., & Quarles, W. (2000). Integrated management of Canada thistle. IPM Practitioner, 22(9), 1-9.
-
Dunsford SJ, Free AJ, Davy AJ, (1998). Acidifying peat as an aid to the reconstruction of lowland heath on arable soil: a field experiment. Journal of Applied Ecology, 35(5):660-672; 25 ref.
-
Edwards, G.R., Bourdot, G.W. and Crawley, M.J. (2000). Influence of herbivory, competition and soil fertility on the abundance of Cirsium arvense in acid grassland. Journal of Applied Ecology, 37, 321–334.
-
Ericsson O. (1997). Control by chemical and biological means. Borss, Sweden: LTs förlag.
-
Fletcher, J. D. (1989). Additional hosts of alfalfa mosaic virus, cucumber mosaic virus, and tobacco mosaic virus in New Zealand. New Zealand journal of crop and horticultural science, 17(4), 361-362.
-
Hill, M. O. (2012). Local frequency as a key to interpreting species occurrence data when recording effort is not known. Methods in Ecology and Evolution, 3(1), 195-205.
-
Hodgson, J. G., & Hunt, R. (1988). Comparative plant ecology: a functional approach to common British species. Unwin Hyman.
-
Holm, L. G., Plucknett, D. L., Pancho, J. V., & Herberger, J. P. (1977). The world’s worst weeds. Distribution and biology. University press of Hawaii.
-
Hongo A. (1989). Weed Res., 29, 7–12.
-
Horie H dan Nemoto M. (1990). Zassou Kenkyu (dalam bahasa Jepang), 35, 340– 345.
-
Kristalova, V., Hejcman, M., Cervena, M. and Pavlu, V. (2011). Effect of nitrogen and phosphorus availability on the emergence, growth and over-wintering of Rumex crispus and Rumex obtusifolius . Grass and Forage Science, 66, 361–369.
-
Jordan, L. S. (1983). Weeds affect citrus growth, physiology, yield, fruit quality. Proceedings of the International Society of Citriculture, 1981. Volume 2., 481-483.
-
Laybourn, R., Kessell, D., Jones, N., Conyers, S., Hallam, C., & Boatman, N. (2013). Review of evidence concerning ragwort impacts, ecology and control options.
-
Mamarot, J., & Roturier, C. (1997). Sensibilité des mauvaises herbes aux herbicides.
-
Manku, R. (1998). Achene variation in bull thistle, Cirsium vulgare (Savi) Ten. Faculty of Graduate Studies, University of Western Ontario.
-
Maskell, L. C., Henrys, P., Pescott, O. L., & Smart, S. M. (2020). Long‐term trends in the distribution, abundance and impact of native “injurious” weeds. Applied Vegetation Science.
-
May, M. J. (2000). Efficiency and selectivity of RR and LL weed control techniques compared to classical weed control systems. 63e Congrès Institut International de Recherches Betteravières, Interlaken, Switzerland, 9-10 fèvrier 2000., 163-170.
-
Miyagi A, Takahashi H, Takahara K, Hirabayashi T, Nishimura Y, Tezuka T, Kawai-Yamada M, dan Uchimiya H. (2010). Metabolomik, 6, 146–155.
-
Mulder, P. P., Klijnstra, M. D., Goselink, R. M., van Vuuren, A. M., Cone, J. W., Stoopen, G., & Hoogenboom, R. L. (2020). Transfer of pyrrolizidine alkaloids from ragwort, common groundsel and viper’s bugloss to milk from dairy cows. Food Additives & Contaminants: Part A, 37(11), 1906-1921.
-
Niggli, U., Nosberger, J. & Lehmann, J., (1993). Effects of nitrogen fertilization and cutting frequency on the competitive ability and the regrowth capacity of Rumex obtusifolius L. in several grass swards. Weed Research, 33, 131–137.
-
Om, H., Dhiman, S. D., Kumar, S., & Kumar, H. (2002). Allelopathic response of Phalaris minor to crop and weed plants in rice–wheat system. Crop Protection, 21(9), 699-705.
-
Oswald, A. K., & Haggar, R. J. (1983). The effects of Rumex obtusifolius on the seasonal yield of two mainly perennial ryegrass swards. Grass and Forage Science, 38(3), 187-191.
-
Panciera RJ, Martin T, Burrows GE, Taylor DS, dan Rice LE, Selai. Dokter hewan. Med. Assoc., 196, 1981–1984 (1990).
-
Parsons, W. T., & Cuthbertson, E. G. (1992). Noxious Weeds of Australia Inkata Press. Melbourne/Sydney.
-
Peel, S. and Hopkins, A. (1980). The incidence of weeds in grasslands. Proceedings 1980 Crop Protection Conference Brighton, 3, 877–890.
-
SAC. (1986). The control of docks. UK, Scottish Agricultural Colleges, Publication No. 56.
-
Sarpe, J., & Mihalcea, G. (1999). Studies on weed control with different herbicides in maize crops [ Zea mays ] in the conditions of the Danube meadow [Romania]. Mededelingen-Faculteit Landbouwkundige en Toegepaste Biologische Wetenschappen Universiteit Gent (Belgium).
-
Suter, M., Siegrist-Maag, S., Connolly, J. and Luscher, A. (2007). Can the occurrence of Senecio jacobaea be influenced by management practice? Weed Research, 47, 262–269.
-
Taylor, K. (2009). Biological flora of the British Isles: Urtica dioica L. Journal of Ecology, 97(6), 1436-1458.
-
Tiley, G.E.D. (2010). Biological Flora of the British Isles: Cirsium arvense (L.) Scop. Journal of Ecology, 98, 938–983.
1 Like